
丂敪惗暘壔峔憿尋媶暘栰偼丄恀妀惗暔偵偍偗傞堚揱巕敪尰惂屼婡峔偺夝柧傪栚巜偟丄斀墳偺拞怱場巕傪恑壔揑崅曐懚場巕偲憐掕偟丄偦傟傜傪撈帺偺庤朄偱婡擻枹抦場巕偲偟偰扨棧偟丄偦偺婡擻夝柧傪峴偭偨忋偱丄惗壔妛丄暘巕惗暔妛丄堚揱妛丄峔憿惗暔妛側偳庤朄偵惂尷傪壛偊偢夝愅傪恑傔丄悽奅傪儕乕僪偡傞撈帺惈偺崅偄條乆側奣擮傪採帵偟偰偄傑偡丅
怴偨側奣擮偺憂弌
怴婯僋儘儅僠儞場巕偺扨棧偲婡擻夝愅
J.Biol.Chem., 272, 30595-30598 (1997)
Genes Cells, 5, 221-233 (2000)
Genes Cells, 5, 251-263 (2000)
Cell, 102, 463-473 (2000)
Proc.Natl.Acad.Sci.U.S.A., 99, 9334-9339 (2002)
Nature Struct.Mol.Biol., 11, 275-283 (2004)
Nature Struct.Mol.Biol., 13, 331-338 (2006)
Proc.Natl.Acad.Sci.U.S.A., 104, 4285-4890 (2007)
僰僋儗僆僜乕儉偺敿曐懚揑暋惢儌僨儖
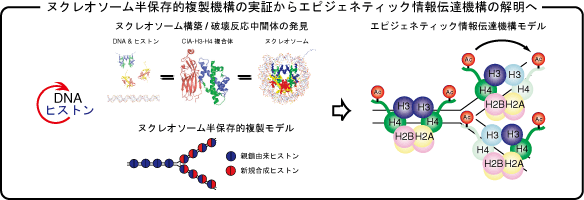
丂恾偼僰僋儗僆僜乕儉偺敿曐懚揑暋惢婡峔偵偮偄偰偺奣擮恾傪帵偟偰偄傑偡丅
丂僰僋儗僆僜乕儉偺暋惢婡峔偱偼僰僋儗僆僜乕儉偺峔惉梫慺僸僗僩儞敧検懱偑柍嶌堊偵暘攝偝傟傞偲嫵壢彂摍偵偼婰弎偝傟偰偄傑偡偑丄摉尋媶幒偱偼30擭棃埨掕偱偁傞偲峫偊傜傟偰偒偨僸僗僩儞(H3-H4)2巐検懱傪擇偮偺僸僗僩儞H3-H4擇検懱偵暘妱偡傞妶惈傪CIA偑桳偡傞偙偲傪帵偟傑偟偨丅偙偺抦尒偐傜丄僰僋儗僆僜乕儉偼DNA摨條丄敿曐懚揑暋惢傪峴偆偙偲偑梊憐偝傟傑偡丅僰僋儗僆僜乕儉偑敿曐懚揑偵暋惢偝傟傞側傜偽丄恾偺塃忋偵帵偡傛偆偵丄僸僗僩儞偺帩偪崌傢偣傞僄僺僕僃僱僥傿僢僋忣曬傕恊嵔僰僋儗僆僜乕儉偐傜柡嵔僰僋儗僆僜乕儉傊嬒摍偵揱払偝傟傞偲峫偊傜傟傑偡丅偙傟傜偺寢壥偵傛傝丄僄僺僕僃僱僥傿僋僗嵟戝偺壽戣傪夝寛偡傞偙偲偵帄傞偺偱偼側偄偐偲峫偊偰偄傑偡丅
Nature, 446, 338-341 (2007)丂
Negotiable border 儌僨儖偺憂弌
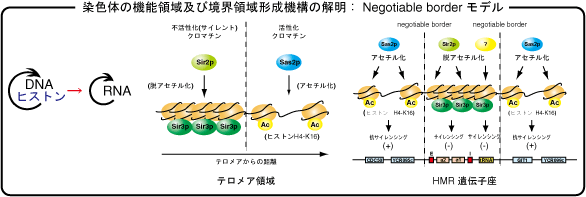
Nature Genet., 32, 370-377 (2002)
Genes Cells, 9, 499-508 (2004)
僸僗僩儞揰曄堎懱偺栐梾揑夝愅傪捠偟偰偺慡妀撪DNA夘嵼斀墳偺夝愅
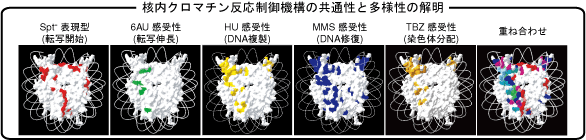
丂僸僗僩儞偼DNA偲偲傕偵愼怓懱偺婎杮峔憿偱偁傞僰僋儗僆僜乕儉傪峔惉偟丄DNA忋偱婲偙傞條乆側斀墳偺梷惂場巕偲偟偰摥偄偰偄傑偡丅偟偨偑偭偰丄揮幨傗DNA暋惢丄DNA廋暅偲偄偭偨DNA忋偱婲偙傞斀墳傪棟夝偡傞偨傔偵偼丄僸僗僩儞丄摿偵斀墳偑婲偙傞夁掱偱條乆側場巕偲憡屳嶌梡偡傞僸僗僩儞偺暘巕昞柺丄偑偦傟傜偺斀墳偵偳偺傛偆偵娭梌偟偰偄傞偐傪柧傜偐偵偡傞偙偲偑昁恵偱偡丅偟偐偟丄1997擭偵僰僋儗僆僜乕儉偺X慄寢徎峔憿偑曬崘偝傟偰偄傞傕偺偺丄僰僋儗僆僜乕儉偺偳偺暘巕昞柺偑偦傟傜偺DNA忋偺斀墳偵娭梌偟偰偄傞偐偼晄柧偱偟偨丅
丂偦偙偱摉尋媶幒偼僰僋儗僆僜乕儉偺暘巕昞柺偵埵抲偡傞320売強偺傾儈僲巁巆婎傂偲偮傂偲偮偵懳偟偰弌夎峺曣偺揰曄堎懱傪嶌惉偟丄揮幨奐巒丄揮幨怢挿丄DNA暋惢丄DNA擇杮嵔愗抐廋暅偵偍偗傞婡擻寚懝傪専弌偡傞専掕傪梡偄偰320屄偺揰曄堎懱偺昞尰宆傪夝愅偟傑偟偨丅偦偺寢壥丄偦傟偧傟偺斀墳宯偱婡擻寚懝傪帵偡揰曄堎懱摨掕偟丄偦傟偧傟偺斀墳宯偵娭梌偡傞僰僋儗僆僜乕儉偺暘巕昞柺傪寛掕偟傑偟偨丅偙傟偵傛傝丄偦傟偧傟偺斀墳宯偵嫟捠偵巊傢傟傞暘巕昞柺偑偁傞堦曽丄偦傟偧傟偺斀墳宯偱巊偄暘偗傜傟偰偄傞暘巕昞柺偑懚嵼偡傞偙偲傪柧傜偐偵偟傑偟偨丅Genes Cells, 12, 13-33 (2007)