
僸僗僩儞僔儍儁儘儞CIA/Asf1偲僸僗僩儞H3, H4偐傜側傞暋崌懱偺峔憿偲婡擻
- 杮榑暥偺攚宨
- 杮榑暥偺撪梕
丂惗暔偺嫟捠婎杮扨埵偱偁傞嵶朎偵偍偄偰丄偦偺峔惉惉暘偺忣曬傪扴偆堚揱巕DNA偼丄僸僗僩儞傪戙昞偲偡傞條乆側抈敀幙偲寢崌偟偰愼怓懱傪宍惉偟偰偄傞丅岝妛尠旝嬀偱娤嶡偱偒傞愼怓懱偼丄堚揱巕偑摥偔偙偲偑偱偒側偄嬅弅偟偨忬懺偱偁傞偑丄庴惛棏偐傜嵶朎偑憹偊側偑傜曄壔偟偰偄偔敪惗抜奒傗丄嵟廔揑偵偼擼丄嬝擏丄寣娗側偳偺慻怐傪宍惉偡傞堎側傞條乆側嵶朎偑惗偠偰偔傞暘壔抜奒偱偼丄愼怓懱忋偺堎側傞埵抲偵偁傞條乆側堚揱巕偑摥偔偨傔偵丄愼怓懱偼偦偺傛偆側嬅弅偟偨枾偺忬懺偐傜丄慳偺忬懺傊偲峔憿偑曄姺偝傟傞昁梫偑偁傞丅
丂1974擭丄愼怓懱偺庡梫峔惉惉暘偱偁傞DNA偲僸僗僩儞偑丄斾妑揑扨弮側婎杮扨埵偱偁傞僰僋儗僆僜乕儉峔憿傪宍惉偟丄DNA忋偵孞傝曉偟暲傫偱偄傞偙偲偑柧傜偐偵偝傟偨丅偙偺尋媶偼揮幨峺慺RNA億儕儊儔乕僛II偺X慄寢徎峔憿夝愅偱嶐擭僲乕儀儖壔妛徿傪庴徿偟偨R.Kornberg偵傛偭偰側偝傟丄偙偺僰僋儗僆僜乕儉峔憿偺敪尒偵傛傝愼怓懱偺峔憿曄姺傪敽偭偰婲偙傞堚揱巕婡擻惂屼偺崻杮揑尨棟偺捛媦偑壜擻偲側偭偨丅
丂偦偺屻丄僰僋儗僆僜乕儉峔憿偑揮幨丄DNA暋惢側偳妀撪斀墳傪梷偊傞摥偒傪帩偮偙偲偑柧傜偐偵側傝丄僰僋儗僆僜乕儉峔憿偺曄姺偑堚揱巕DNA偺帩偮忣曬傪堷偒弌偡偐斲偐傪寛傔傞丄偡側傢偪堚揱巕敪尰偺ON/OFF偺僗僀僢僠偺崻杮揑婎斦偵側偭偰偄傞偙偲偑柧傜偐偵偝傟偰偒偨丅廬偭偰丄愼怓懱傪峔惉偡傞婎杮扨埵偱偁傞僰僋儗僆僜乕儉峔憿偺曄姺婡峔傪柧傜偐偵偡傞偙偲偼丄愼怓懱偺帩偮慡偰偺摥偒媦傃偦偺儊僇僯僘儉傪棟夝偡傞忋偱丄嵟傕廳梫側壽戣偲側偭偨
丂丂
丂1997擭偵K.Luger, T.Richmond傜偵傛偭偰僰僋儗僆僜乕儉偺X慄寢徎峔憿夝愅偑側偝傟丄僸僗僩儞H2A, H2B, H3, H4偑2暘巕偢偮廤傑偭偨僸僗僩儞敧検懱偺廃傝傪丄146墫婎懳偐傜側傞DNA偑1.75廃姫偒偮偄偰偄傞偙偲偑柧傜偐偲側偭偨乮恾侾乯丅偟偐偟側偑傜丄偙偺僰僋儗僆僜乕儉峔憿偑偳偺傛偆偵宍惉偝傟丄昁梫偵墳偠偰偳偺傛偆偵攋夡偝傟傞偺偐偼尰嵼偵帄傞傑偱埶慠偲偟偰撲偵曪傑傟偨傑傑偱偁偭偨丅杮榑暥偱偼丄僸僗僩儞僔儍儁儘儞CIA偲僸僗僩儞H3, H4偐傜側傞暋崌懱偺X慄寢徎峔憿夝愅偺寢壥乮恾俀乯丄僰僋儗僆僜乕儉撪偱僸僗僩儞H3摨巑偺憡屳嶌梡偵梡偄傜傟傞暘巕昞柺傪CIA偑棙梡偟偰僸僗僩儞H3偲憡屳嶌梡偡傞偙偲偵傛傝丄CIA-僸僗僩儞H3-H4暋崌懱傪宍惉偟偰偄傞偙偲丄偡側傢偪僰僋儗僆僜乕儉峔憿曄姺斀墳偺拞娫忬懺傪庢偭偰偄傞偙偲傪帵嵈偡傞抦尒傪摼傞偙偲偵惉岟偟偨乮恾俁乯丅偙偺偙偲偵傛傝丄崱傑偱慡偔抦傞偙偲偺偱偒側偐偭偨僰僋儗僆僜乕儉峔憿曄姺惂屼婡峔丄峏偵偼愼怓懱峔憿曄姺惂屼婡峔偵偮偄偰丄夝柧偺抂弿偲側傞儌僨儖傪採帵偡傞偙偲偑偱偒偨丅
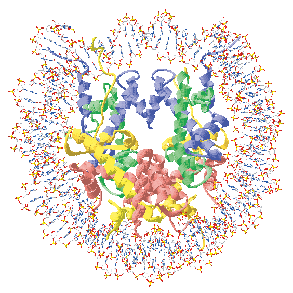
恾侾丏僰僋儗僆僜乕儉棫懱峔憿恾丅恾拞偵偍偄偰惵偑僸僗僩儞H3丄椢偑僸僗僩儞H4丄墿怓偑僸僗僩儞H2A丄愒偑H2B傪偁傜傢偡丅H3, H4, H2A, H2B偑2暘巕偢偮廤傑偭偨僸僗僩儞敧検懱偺夞傝傪DNA偑姫偒偮偄偰偄傞丅偙偺峔憿拞偱偼丄H3-H3乫憡屳嶌梡偑峔憿堐帩偵廳梫側摥偒傪扴偭偰偄傞丅
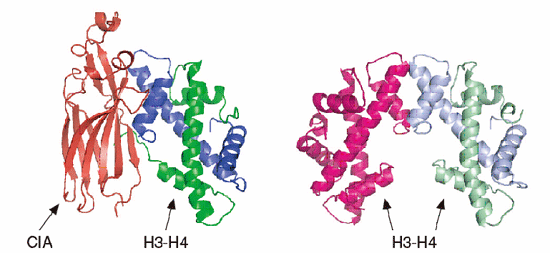
恾俀丏CIA-僸僗僩儞H3-H4暋崌懱峔憿偲丄僰僋儗僆僜乕儉峔憿拞偵偍偗傞僸僗僩儞H3-H4擇検懱娫偺憡屳嶌梡偺斾妑丅僸僗僩儞H3-H3乫憡屳嶌梡晹埵偵CIA偑憡屳嶌梡偟丄CIA偑僸僗僩儞摨巑偺憡屳嶌梡傪杊偄偱偄傞偙偲偑傢偐傞丅
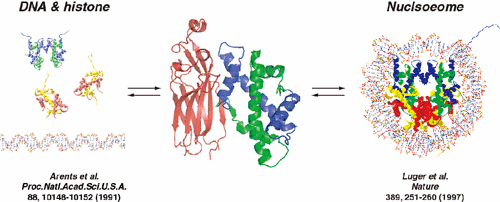
恾俁丏CIA-僸僗僩儞H3-H4暋崌懱偼僰僋儗僆僜乕儉峔憿偺宍惉丄攋夡斀墳偵偍偗傞拞娫懱偱偁傞偲峫偊傜傟傞丅
丂杮榑暥偱偼偦傟埲奜偵傕廳梫側揰傪柧傜偐偵偡傞偙偲偑偱偒偨丅僸僗僩儞偺暋悢偺傾儈僲巁巆婎偼條乆側奜奅偺娐嫬偺曄壔傗嵶朎偺敪惗丒暘壔忬懺偵墳偠偰傾僙僠儖壔傗儊僠儖壔側偳條乆側壔妛廋忺傪庴偗偰偄傞丅偙偺傛偆偵偟偰寛掕偝傟偨愼怓懱偺奺椞堟偵偍偗傞僸僗僩儞偺壔妛廋忺僷僞乕儞偑堚揱巕敪尰僷僞乕儞傪寛掕偟偰偄傞偙偲偑嬤擭媫懍偵懡偔偺椺偱柧傜偐偵側偭偰偒偨丅敪惗丒暘壔偵敽偄丄摨偠堚揱巕敪尰僷僞乕儞傪桳偡傞嵶朎偑憹偊傞偙偲偐傜丄偙偺堚揱巕敪尰僷僞乕儞偵娭偡傞忣曬偑墫婎攝楍偵埶傜偢恊嵶朎偐傜柡嵶朎傊揱払偝傟偰偄傞偙偲偑帵嵈偝傟偰偄傞丅偙傟偼堚揱巕偺墫婎攝楍偵傛偭偰寛傑傞僕僃僱僥傿僋僗偵懳偟丄僄僺僕僃僱僥傿僋僗偲屇偽傟偰偍傝丄僸僩傗儅僂僗僎僲儉偺慡墫婎攝楍偑寛掕偝傟偨億僗僩僎僲儉帪戙偵擖偭偨尰嵼傕丄埶慠偲偟偰撲偵曪傑傟偨嵟廳梫壽戣偲側偭偰偄傞丅偡側傢偪丄偙偺僸僗僩儞偺壔妛廋忺僷僞乕儞偑偳偺傛偆偵恊嵶朎偐傜柡嵶朎傊偲庴偗宲偑傟偰偄偔偺偐丄偑戝偒側栤戣偲側偭偰偄傞丅廬偭偰丄恊嵶朎偲摨偠堚揱巕敪尰僷僞乕儞傪帩偮2偮偺柡嵶朎傪嶌傞偵偼丄DNA暋惢偵敽偭偰恊DNA嵔偑帩偮僰僋儗僆僜乕儉偵偍偗傞擟堄偺僸僗僩儞偺壔妛廋忺僷僞乕儞傪堐帩偟側偑傜丄2杮偺柡DNA嵔偦傟偧傟偵怴偨側僰僋儗僆僜乕儉傪宍惉偟側偗傟偽側傜側偄偲峫偊傜傟傞丅偟偐偟丄偙偺婡峔傪峫偊傞偵偼戝偒側忈暻偑懚嵼偟偰偄偨丅
丂僰僋儗僆僜乕儉偼僸僗僩儞H2A, H2B, H3, H4偑2暘巕偢偮廤傑偭偨峔憿偱偁傞偨傔丄僸僗僩儞偺壔妛廋忺忬懺傪堐帩偡傞偨傔偺嵟傕扨弮側儊僇僯僘儉偲偟偰峫偊傜傟傞偺偼丄恊DNA嵔偑帩偮僰僋儗僆僜乕儉偐傜僸僗僩儞H2A, H2B, H3, H4傪奺乆1暘巕偢偮2偮偺柡DNA嵔偵暘攝偡傞曽朄偱偁傞乮恾係乯丅偙偺儊僇僯僘儉偼丄柡DNA嵔偺帩偮僰僋儗僆僜乕儉偑恊DNA嵔桼棃偺僸僗僩儞偲怴婯偵崌惉偝傟偨僸僗僩儞偐傜側偭偰偄傞偲偄偆揰偱丄DNA偺敿曐懚揑暋惢偺儊僇僯僘儉乮恾俆乯偵懳墳偡傞偲傕偄偊傞丅
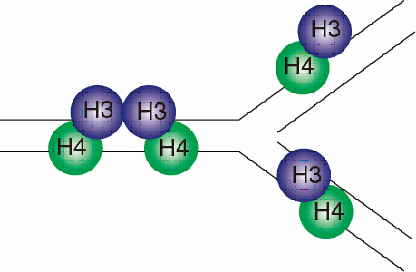
恾係丏僰僋儗僆僜乕儉偺暋惢傪峫偊傞忋偱丄嵟傕扨弮側僸僗僩儞暘攝偺儊僇僯僘儉丅
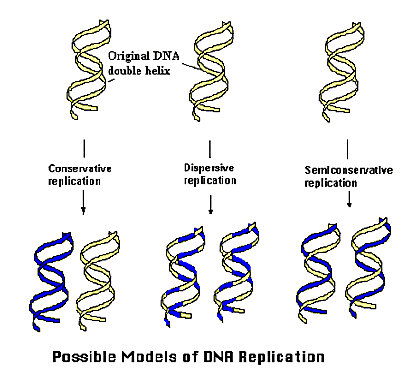
恾俆丏DNA偺暋惢婡峔偺儌僨儖丅1953擭偵DNA偺峔憿儌僨儖偑J.Watson偲F.Crick偵傛偭偰採彞偝傟傞偙偲偱丄DNA暋惢婡峔偲偟偰慡曐懚揑乮Conservative乯丄暘嶶揑暋惢乮Dispersive乯丄敿曐懚揑暋惢乮Semi-conservative乯偺儌僨儖偑峫偊弌偝傟丄1958擭偵M.Meselson偲F.Stahl偵傛偭偰敿曐懚揑暋惢儊僇僯僘儉偑惓偟偄偙偲偑徹柧偝傟偨丅
丂偲偙傠偑丄僸僗僩儞H3, H4偼僸僗僩儞H3-H3乫娫偺嫮偄憡屳嶌梡傪婎杮偲偟偨巐検懱傪宍惉偟偰偄傞偲挿傜偔峫偊傜傟偰偄偨偨傔丄僰僋儗僆僜乕儉偺敿曐懚揑暋惢傪峫偊傞偙偲偵偼柍棟偑偁偭偨丅挿偄娫怣偠傜傟偰偒偨偦偺峫偊傪曄峏偡傞偵偼僸僗僩儞(H3-H4)2巐検懱傪攋夡偡傞壗傜偐偺場巕偑偁傞偙偲傪徹柧偟丄偟偐傕僰僋儗僆僜乕儉偺敿曐懚揑暋惢偵偍偗傞嶌梡儊僇僯僘儉傪柧傜偐偵偟側偗傟偽側傜側偄乮恾俇乯丅峏偵丄恊DNA嵔偺帩偮僰僋儗僆僜乕儉偵偍偗傞擟堄偺僸僗僩儞偺壔妛廋忺僷僞乕儞傪柡DNA嵔偺帩偮僰僋儗僆僜乕儉傊偲揱払偡傞偨傔偵偼丄僸僗僩儞壔妛廋忺偵懳偟偰傑偭偨偔塭嬁傪梌偊偢偵丄僰僋儗僆僜乕儉峔憿偺宍惉丄攋夡傪峴偆偙偲偑壜擻側嶌梡儊僇僯僘儉偱偁傞偙偲偑昁梫晄壜寚偱偁偭偨丅
恾俇丏僰僋儗僆僜乕儉偺敿曐懚揑暋惢偵偼丄僸僗僩儞(H3-H4)2巐検懱傪攋夡偡傞場巕偑昁梫偱偁傞丅
丂杮榑暥偱偼丄摉尋媶幒偱悽奅偵愭嬱偗偰扨棧丄婡擻夝愅傪恑傔偰偒偨僸僗僩儞僔儍儁儘儞CIA偵忋婰偺嶌梡儊僇僯僘儉傪惉傝棫偨偣傞惈幙偑偁傞偙偲傪柧傜偐偵偟偰偄傞乮恾俀,俇,俈乯丅偙偺寢壥偼丄1974擭偺R.Kornberg偵傛傞僰僋儗僆僜乕儉峔憿偺敪尒丄1997擭偺K.Luger, T.Richmond傜偵傛傞僰僋儗僆僜乕儉偺棫懱峔憿夝柧埲棃丄弶傔偰僰僋儗僆僜乕儉峔憿曄姺婡峔儌僨儖傪採彞偟偆傞夝愅傪堄枴偟偰偍傝丄愼怓懱偺峔憿曄姺惂屼偺夝柧偵戝偒偔婑梌偟偆傞傕偺偱偁傞丅傑偨丄1953擭偺J.Watson偲F.Crick偺DNA偺敿曐懚揑暋惢儌僨儖偵懳偟偰偺DNA-僸僗僩儞偐傜側傞僰僋儗僆僜乕儉峔憿偺敿曐懚揑暋惢儌僨儖傪採彞偡傞傕偺偱偁傝乮恾俉乯丄愼怓懱偺婡擻曄姺惂屼丄偡側傢偪僄僺僕僃僱僥傿僋僗惂屼偺巇慻傒傪夝柧偡傞戞堦曕偺惉壥偱偁傞偲偄偊傞乮恾俈,俋乯丅
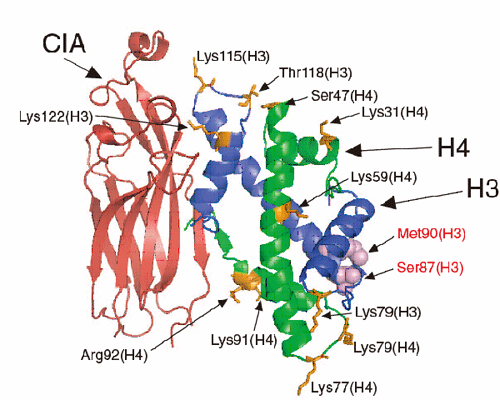
恾俈丏崱夞偺峔憿夝愅偵傛傝丄CIA-僸僗僩儞H3-H4暋崌懱峔憿拞偱僸僗僩儞H3媦傃H4偺壔妛廋忺巆婎偑業弌偝傟偰偄傞偙偲偑暘偐偭偨丅偙偺偙偲偐傜丄CIA偼僸僗僩儞H3媦傃H4偺壔妛廋忺偵懳偟偰塭嬁傪梌偊偢偵丄僰僋儗僆僜乕儉峔憿偺宍惉丄攋夡傪峴偆偙偲偑壜擻偱偁傞偙偲偑帵嵈偝傟偨丅
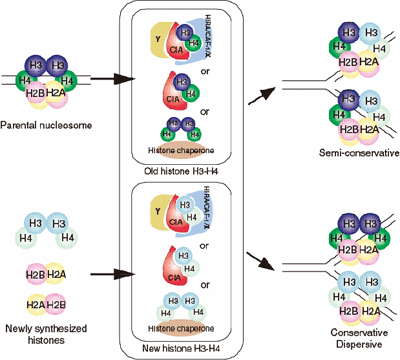
恾俉丏崱夞偺峔憿夝愅偵傛傝丄僰僋儗僆僜乕儉傕敿曐懚揑偵暋惢偝傟傞壜擻惈偑帵偝傟偨丅
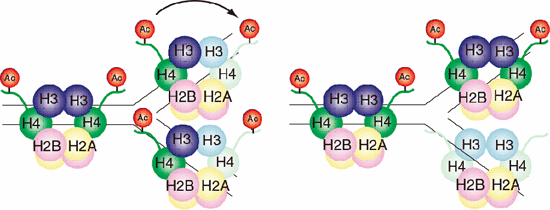
恾俋丏恊DNA嵔偺帩偮僰僋儗僆僜乕儉偵偍偗傞擟堄偺僸僗僩儞偺壔妛廋忺僷僞乕儞偑柡DNA嵔偺帩偮僰僋儗僆僜乕儉傊偲揱払偝傟傞丅